
Efecto de la lateralización cerebral en el comportamiento de aprendizaje de Poecilia reticulata
Effect of Cerebral Lateralization on the Learning Behavior of Poecilia reticulata
Arianis Mojica Lezcano1
https://orcid.org/0009-0007-2663-764X
Reggie Guerra Montenegro2,3
https://orcid.org/0000-0001-8471-2862
Reynaldo Vargas Vargas2,3
https://orcid.org/0000-0002-5420-9761
1 Colegio Secundario de Alanje. Proyecto Jóvenes Científicos 2022 SENACYT
2 Universidad de Panamá, Facultad de Ciencias Agropecuarias, Departamento de Zootecnia
3 Sistema Nacional de Investigación de la Secretaría Nacional de Ciencia, Tecnología e Innovación (SNI-SENACYT)
Autor correspondiente: reggie.guerra@up.ac.pa
Enviado el: 10 de octubre de 2023 Aceptado el: 13 de diciembre de 2023
https://doi.org/10.59722/rcvn.v1i2.704
Resumen
Lateralización cerebral se refiere al análisis preferencial de tipos particulares de información y procesos en un hemisferio cerebral específico y se cree que esta influye en un mayor rendimiento cognitivo. El objetivo del proyecto fue evaluar el efecto de la lateralización cerebral en el comportamiento de aprendizaje de Poecilia reticulata. Se separaron 60 individuos en tres fenotipos derecho, izquierdo y no lateralizado. Se seleccionaron ocho individuos lateralizados derechos e izquierdos y seis no lateralizados para la prueba cognitiva de discriminación visual y aprendizaje inverso. Los peces lateralizados representaron el 88 % de la población (χ12=115.5; p<0.001). No se encontró diferencias significativas en el porcentaje de individuos que se orientó hacia la izquierda o derecha (χ12=1.28; p=0.25). No se encontró diferencias significativas en la prueba de discriminación visual entre los tratamientos (K-W=3.97; p=0.13). Se observaron diferencias significativas entre las repeticiones (K-W=8.00; p=0.04). La interacción tratamiento por repeticiones no mostró diferencias significativas (K-W=14.55; p=0.20). En la prueba de aprendizaje inverso los fenotipos mostraron diferencias significativas en las latencias de entrada al nuevo ambiente (K-W=6.65; p=0.03). No se encontró diferencias significativas en las repeticiones (K-W=0.99; p=0.80), ni en la interacción tratamiento por repeticiones (K-W=10.25; p=0.50). La población estudiada muestra más del 80 % de lateralización cerebral. El porcentaje de individuos lateralizados izquierdos o derechos fue similar. El nivel cognitivo fue similar entre los fenotipos en la prueba de discriminación visual. En la prueba de aprendizaje inverso el fenotipo no lateralizado mostró mayor flexibilidad que los fenotipos lateralizados.
Palabras clave
Aprendizaje, fenotipos, latencia, pruebas cognitivas.
Abstract
Cerebral lateralization refers to the preferential analysis of specific types of information and processes in a specific cerebral hemisphere, believed to influence higher cognitive performance. The project aimed
to assess the effect of cerebral lateralization on the learning behaviour of Poecilia reticulata. Sixty individuals were categorized into three phenotypes: right-lateralized, left-lateralized, and non-lateralized. Eight right-lateralized, eight left-lateralized, and six non-lateralized individuals were selected
for the visual discrimination and reverse learning cognitive test. Lateralized fish represented 88% of the population (χ12=115.5; p<0.001). No significant differences were found in the percentage of individuals orienting to the left or right (χ12=1.28; p=0.25). No significant differences were observed between treatments in the visual discrimination test (K-W=3.97; p=0.13). Significant differences were noted between repetitions (K-W=8.00; p=0.04). The treatment by repetitions interaction did not show significant differences (K-W=14.55; p=0.20). In the reverse learning test, phenotypes exhibited significant differences in entry latencies to the new environment (K-W=6.65; p=0.03). No significant differences were found in repetitions (K-W=0.99; p=0.80), nor the treatment by repetitions interaction (K-W=10.25; p=0.50). The studied population demonstrates over 80% cerebral lateralization. The percentage of left or right lateralized individuals was similar. Cognitive levels were comparable among phenotypes in the visual discrimination test. In the reverse learning test, the non-lateralized phenotype showed greater flexibility than the lateralized phenotype.
Keywords
Latency, learning, cognitive tests, phenotypes.
Introducción
La lateralización cerebral describe un procesamiento asimétrico de funciones cognitivas entre los dos hemisferios cerebrales y, con mayor frecuencia, se muestra abiertamente como un sesgo conductual, como la "dominancia". El sesgo conductual resultante se conoce como lateralidad (Berlinghieri et al. 2021).
La naturaleza generalizada de la lateralidad entre los taxones de vertebrados sugiere que se trata de una antigua innovación evolutiva y probablemente conduce al uso más eficiente del costoso tejido neural para obtener la máxima potencia de procesamiento (Magat & Brown, 2009).
La existencia de la variabilidad en las respuestas de comportamiento sugiere que la lateralidad podría ocurrir bajo la selección de equilibrio, en la que puede haber contextos en los que la lateralidad no proporciona un beneficio de aptitud o incluso puede ser costoso (Dadda et al. 2009). Muchas investigaciones se han enfocado en la comprensión de los costos y beneficios de la lateralización cerebral, con estudios de campo y de laboratorio que demuestran capacidades cognitivas mejores en individuos lateralizados (Dadda et al. 2009; Irving & Brown, 2013).
El hecho de que la lateralidad sea un rasgo conductual tan generalizado sugiere fuertemente que tiene ventajas de aptitud física, muchas de las cuales parecen estar relacionadas con la función cognitiva. Generalmente se cree que el desarrollo y evolución de la lateralización es un mecanismo ventajoso que mejora la capacidad cognitiva y la multitarea al permitir el procesamiento de múltiples estímulos externos (Brown, 2005).
Varios estudios (Ferrari et al., 2017; Roche et al., 2013; Roche et al., 2020) también informan un alto grado de variación individual en la fuerza de la lateralización en individuos medidos repetidamente en diferentes contextos, lo que sugiere que este rasgo puede ser más lábil de lo que se creía anteriormente.
Debido a las metodologías simples que se pueden emplear para la evaluación de la lateralidad y el impacto de varios factores ecológicos y de estrés en la lateralidad, existe cada vez mayor interés en el uso de la lateralidad para evaluar el bienestar de los peces (Berlinghieri et al. 2021). Por tanto, se planteó como objetivo evaluar el efecto de la lateralización cerebral en el comportamiento de aprendizaje de Poecilia reticulata.
Materiales y Métodos
Sesenta peces Poecilia reticulata adultos fueron adquiridos en un proveedor local. Se estabularon en un tanque de 60 litros, con ambiente controlado (condiciones estándar). Se alimentaron hasta la saciedad dos veces al día, en la mañana con un pienso seco y en la tarde con un pienso húmedo de acuerdo con las especificaciones del fabricante tanto durante el periodo de aclimatación como durante la fase experimental.
Para la prueba de lateralización se realizó una adaptación a un aparato de prueba propuesto por Bisazza et al. (1997), utilizando un tanque (47cm de largo y 33cm de ancho) con una pista en el centro (10 cm de ancho y 30 cm de largo) que permitía a los peces observar una barrera en la parte final de la misma y elegir la dirección derecha o izquierda de salida (figura 1).

Figura 1.
Aparato para prueba de lateralización cerebral
Durante cada prueba, se empujaba suavemente a los peces con un par de redes de pesca en el punto de inicio de la pista. El pez se desplazó a lo largo de la pista hasta que se enfrentó a la barrera. Se registró la dirección derecha o izquierda de la respuesta de desvío en una serie de 10 ensayos consecutivos. La clasificación en no lateralizado, lateralizado izquierdo o derecho se realizó utilizando un índice en donde (derecho - izquierdo) / (derecho + izquierdo) x 100 = índice de lateralidad (Bisazza et al. 1997), en donde aquellos individuos con porcentajes positivos se desvían hacia la derecha fijando el objetivo con el ojo izquierdo y aquellos con valores negativos hacia la izquierda fijando el objetivo con el ojo derecho. La clasificación se verificó siguiendo la metodología propuesta por Amira et al. (2018) y Reddon et al. (2008).
En el caso de la prueba de discriminación visual y aprendizaje inverso se eligieron ocho peces de los grupos lateralizados (derecho e izquierdo) y seis de los no lateralizados. Cada una de las pruebas se aplicó de forma individual por cuatro días seguido y ambas pruebas de forma consecutiva.
En la prueba de discriminación visual el tanque de prueba fue similar al empleado en la prueba de selección de fenotipos de lateralización cerebral. Se adaptó una barrera trasparente que permitía al pez observar el nuevo ambiente, en cada uno de los lados de la barrera trasparente se colocó una figura geométrica, al lado de una de las figuras geométricas se habilitó una puerta para permitir la entrada al nuevo ambiente (figura 2).
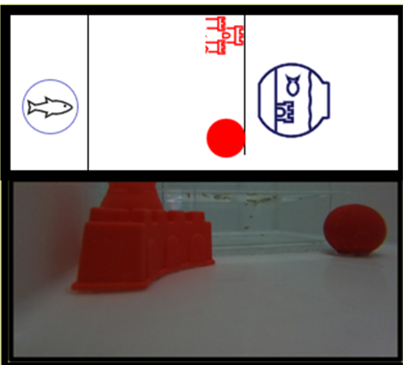
Figura 2.
Aparato para prueba de aprendizaje y aprendizaje inverso.
Esta prueba fue adaptada de Hirata et al. (2016), y consistió en colocar un pez en el extremo inicial del tanque y evaluar el tiempo en segundos (latencia) que demora en alcanzar el ambiente nuevo ubicado detrás de barrera transparente. En el caso del estudio se utilizó como estímulo un grupo de peces en una pecera plástica de siete litros la cual fue ubicada en el extremo del tanque detrás de la barrera trasparente (Versace et al. 2007). La prueba fue aplicada durante cuatro días consecutivos. En la prueba de aprendizaje inverso el aparato de prueba y la variable evaluada fueron similares a las descritas en la prueba de discriminación visual. El procedimiento se modificó realizando cambio de posición de las figuras geométricas y la puesta para ingresar al ambiente nuevo.
Los datos obtenidos se tabularon en una hoja de cálculo de Microsoft Excel® 2021. El análisis estadístico, se realizó con R 2.12 (R-Developement-Core-Team 2021), Statistica versión 10 (StatSoftV10) y los gráficos mediante Graph Pad Prism V.8.0.0 (San Diego, CA, USA). La distribución porcentual de fenotipos de lateralización cerebral fue evaluada con la prueba de Chi Cuadrado. Los valores de latencia para ingresar en ambiente nuevo en la prueba de discriminación visual y aprendizaje inverso se les verificó la normalidad mediante la prueba de Shapiro-Wilk, y posteriormente, la homocedasticidad de varianzas con la prueba de Levene. La variable mostró una distribución no normal y fue analizada utilizando la prueba no paramétrica de Kruskal-Wallis y corregida con la prueba de Dunn´s. Los resultados se expresan como media ± EE (Error estándar). Se empleó un nivel de significancia de p<0.05.
Resultados
Prueba de lateralización
Los peces lateralizados (figura 3) representaron el 88 % de la población estudiada (χ12=115.5; p<0.001). El porcentaje de individuos que se orientó hacia la izquierda o derecha no mostraron diferencias significativas (χ12=1.28; p=0.25, figura 4). Existen diferencias significativas en la distribución porcentual de orientaciones hacia la izquierda o derecha en cada uno de los grupos lateralizados (χ12=54.43; p<0.001, figura 5).

Figura 3.
Distribución porcentual de animales lateralizados (L) y no lateralizados (NL) en la población estudiada.

Figura 4.
Distribución porcentual de individuos lateralizados izquierdo (LI) y derecho (LD) del total de la población lateralizada.
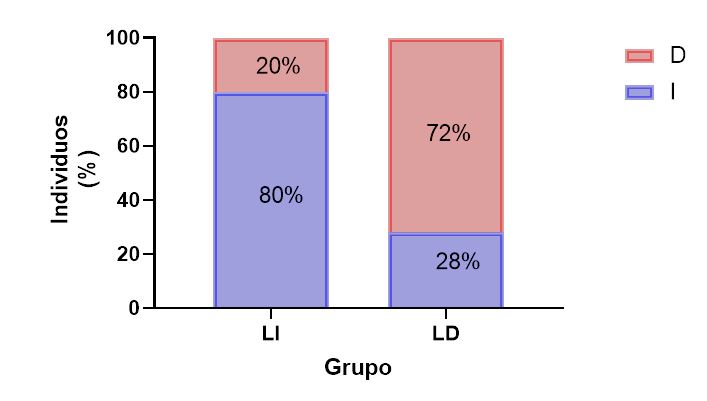
Figura 5.
Distribución porcentual de las orientaciones izquierda o derecha, tomadas por los individuos durante la prueba de lateralización de cada uno de los grupos lateralizados.
Prueba de discriminación visual de aprendizaje
La figura 6, muestra que existen diferencias significativas en las repeticiones en la prueba de discriminación visual (K-W=6.65; p=0.03). No se encontraron diferencias significativas en la prueba de discriminación visual entre los tratamientos (K-W=3.97; p=0.13). La interacción tratamiento por repeticiones no mostró diferencias significativas (K-W=14.55; p=0.20).
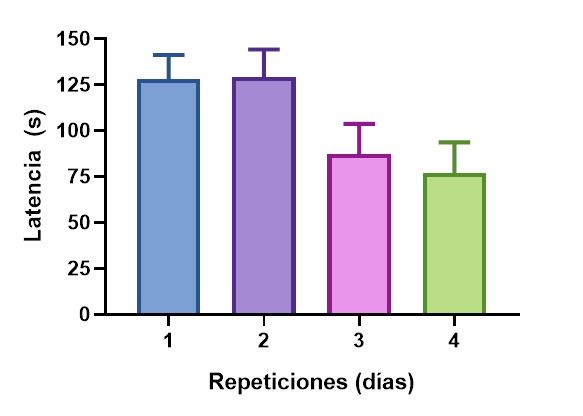
Figura 6.
Latencia en segundos (s) durante los cuatro días que se aplicó la prueba de aprendizaje visual.
Prueba de aprendizaje inverso
En la prueba de aprendizaje inverso los fenotipos (figura 7) mostraron diferencias significativas en las latencias de entrada al nuevo ambiente (K-W=6.65; p=0.03). No se encontraron diferencias significativas en las repeticiones (K-W=0.99; p=0.80), ni en la interacción tratamiento por repeticiones (K-W=10.25; p=0.50).
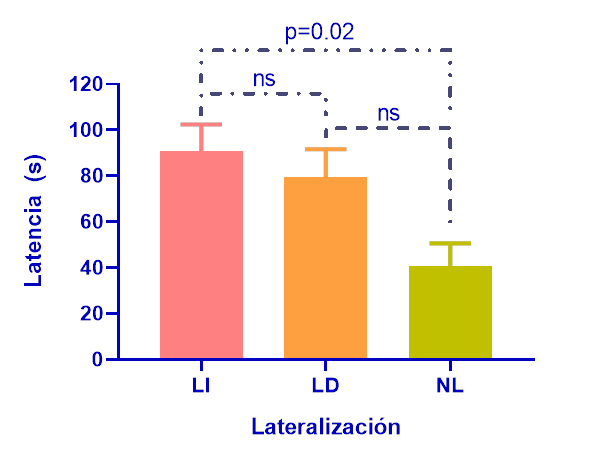
Figura 7.
Latencia en segundos en la prueba de aprendizaje inverso entre individuos lateralizados izquierdos (LI) lateralizados derechos (LD) y no lateralizados (NL). Valores de p<0.05 son significativos.
Discusión
Los resultados de estudio muestran que un 88% de la población estudiada presentaba lateralización, lo cual no es sorprendente si tomamos en cuenta que la lateralización cerebral permite ventajas competitivas en entornos que muestran desafíos ambientales. Estos resultados pueden estar en línea con lo propuesto por (Dadda & Bisazza, 2006) indicando que los peces lateralizados pueden realizar un seguimiento del depredador con un ojo y procesar esta información en el hemisferio opuesto, lo que
permite el procesamiento de otras tareas, como la formación de bancos o la búsqueda de alimento, con el hemisferio restante.
Los individuos no lateralizados, en comparación, tienden a cambiar entre tareas y usan ambos ojos. En ese mismo contexto se indicó que en Poecilia reticulata la lateralización aumenta las habilidades de los individuos para la formación de grupos (Gatto et al. 2019). En la prueba de cognición de discriminación visual no observamos diferencias entre los grupos lateralizados y no lateralizados, sin embargo, observamos una reducción gradual de la latencia para ingresar en un ambiente nuevo que puede estar asociada a un proceso de habituación (Murphy & Arkins, 2007). En este sentido se ha demostrado que los factores estresantes repetidos desensibilizan a los peces y moderan las respuestas neuroendocrinas a factores estresantes similares lo que comúnmente se conoce como habituación (Staven et al. 2019). La habituación, medida como respuestas neuroendocrinas desensibilizadas, por ejemplo, reducción de la secreción de cortisol durante interacciones depredadoras repetidas, se ha demostrado previamente en especies modelo de pez cebra Danio rerio Hamilton, 1822 (Barcellos et al. 2010).
Respecto a las pruebas de aprendizaje inverso detectó variabilidad en la capacidad cognitiva entre los grupos lateralizados izquierdo y los no lateralizados. Los peces no lateralizados mostraron menor latencia en ingresar al ambiente nuevo que los peces lateralizados izquierdo. Al realizar la inversión de la puerta para ingresar al ambiente nuevo, ésta quedaba en el lado opuesto al ojo mayormente utilizado por los peces lateralizados izquierdo lo cual podría explicar la mayor latencia en este grupo. Otra posible explicación es que nosotros utilizamos como estimulo otros peces y de acuerdo con un modelo propuesto por (Bisazza et al. 1997), según el cual los peces poecílidos tienden a usar el ojo derecho durante la fijación de estímulos biológicamente relevantes, como una hembra o un depredador, en nuestro estudio la ubicación de la entrada en la prueba de aprendizaje inverso afectaría aquellos peces que dependen más del ojo izquierdo.
La menor latencia para ingresar a un ambiente nuevo en los peces no lateralizado puede deberse a que cuando existe incremento en los niveles de miedo la información es procesada por un solo hemisferio (Brown & Bibost, 2014), en nuestro caso al invertir la entrada al nuevo ambiente incremento los niveles de estrés de los animales lo que pudo darles ventajas a los peces no lateralizados. En este contexto (Dadda et al. 2009), reportaron que en una especie de pez individuos no lateralizados superan a los individuos lateralizados tanto en la elección de cardúmenes bilaterales que los individuos lateralizados.
Conclusión
Los resultados indican que la lateralización cerebral tiene un efecto en el aprendizaje, los peces lateralizados izquierdos mostraron mayor latencia para entrar en un ambiente nuevo en la prueba de reversión, sugiriendo una menor capacidad de memoria espacial para evaluar el nuevo entorno lo cual apoya resultados de previos estudios.
Referencias
Amira, A. G., Gareth, P. P., Jashim, U., Eloise, R., Harriet, D., & Clive, J. C. P. (2018). A forced lateralisation test for dairy cows and its relation to their behaviour. Applied Animal Behaviour Science, 207, 8–19. https://doi.org/10.1016/j.applanim.2018.06.008
Barcellos, L. J. G., Ritter, F., Kreutz, L. C., & Cericato, L. (2010). Can zebrafish Danio rerio learn about predation risk? The effect of a previous experience on the cortisol response in subsequent encounters with a predator. Journal of Fish Biology, 76(4), 1032–1038. https://doi.org/10.1111/j.1095-8649.2010.02542.x
Berlinghieri, F., Panizzon, P., Penry-Williams, I. L., & Brown, C. (2021). Laterality and fish welfare - A review. Applied Animal Behaviour Science, 236, 105239. https://doi.org/10.1016/j.applanim.2021.105239
Bisazza, A., Pignatti, R., & Vallortigara, G. (1997). Laterality in detour behaviour: Interspecific variation in poeciliid fish. Animal Behaviour, 54(5), 1273–1281. https://doi.org/10.1006/anbe.1997.0522
Brown, C. (2005). Cerebral lateralisation, “social constraints”, and coordinated anti-predator responses. Behavioral and Brain Sciences, 28(4), 591–592. https://doi.org/10.1017/S0140525X05240104
Brown, C., & Bibost, A. L. (2014). Laterality is linked to personality in the black-lined rainbowfish, Melanotaenia nigrans. Behavioral Ecology and Sociobiology, 68(6), 999–1005. https://doi.org/10.1007/s00265-014-1712-0
Dadda, M., & Bisazza, A. (2006). Does brain asymmetry allow efficient performance of simultaneous tasks? Animal Behaviour, 72(3), 523–529. https://doi.org/10.1016/j.anbehav.2005.10.019
Dadda, M., Zandonà, E., Agrillo, C., & Bisazza, A. (2009). The costs of hemispheric specialization in a fish. Proceedings of the Royal Society B: Biological Sciences, 276(1677), 4399–4407.
https://doi.org/10.1098/rspb.2009.1406
Ferrari, M. C. O., McCormick, M. I., Mitchell, M. D., Allan, B. J. M., Gonçalves, E. J., & Chivers, D. P. (2017). Daily variation in behavioural lateralization is linked to predation stress in a coral reef fish. Animal Behaviour, 133, 189–193. https://doi.org/10.1016/j.anbehav.2017.09.020
Gatto, E., Agrillo, C., Brown, C., & Dadda, M. (2019). Individual differences in numerical skills are influenced by brain lateralization in guppies (Poecilia reticulata). Intelligence, 74(April), 12–17. https://doi.org/10.1016/j.intell.2019.04.004
Hirata, M., Tomita, C., & Yamada, K. (2016). Use of a maze test to assess spatial learning and memory in
cattle: Can cattle traverse a complex maze?. Applied Animal Behaviour Science, 180, 18-25.
Irving, E., & Brown, C. (2013). Examining the link between personality and laterality in a feral guppy poecilia reticulata population. Journal of Fish Biology, 83(2), 311–325. https://doi.org/10.1111/jfb.12165
Magat, M., & Brown, C. (2009). Laterality enhances cognition in Australian parrots. Proceedings of the
Royal Society of London B: Biological Sciences, 276(1676), 4155-4162.
Murphy, J., & Arkins, S. (2007). Equine learning behaviour. Behavioural Processes, 76(1), 1–13. https://doi.org/10.1016/j.beproc.2006.06.009
Reddon, A. R., Hurd, P. L., Reddon, A. R., & Hurd, P. L. (2008). Aggression , sex and individual differences in cerebral lateralization in a cichlid fish Aggression , sex and individual differences in cerebral lateralization in a cichlid fish, 338–340. https://doi.org/10.1098/rsbl.2008.0206
Roche, D. G., Binning, S. A., Strong, L. E., Davies, J. N., & Jennions, M. D. (2013). Increased behavioural lateralization in parasitized coral reef fish. Behavioral Ecology and Sociobiology, 67(8), 1339–1344. https://doi.org/10.1007/s00265-013-1562-1
Roche, D. G., Amcoff, M., Morgan, R., Sundin, J., Andreassen, A. H., Finnøen, M. H., Lawrence, M. J., Henderson, E., Norin, T., Speers-Roesch, B., Brown, C., Clark, T. D., Bshary, R., Leung, B., Jutfelt, F., & Binning, S. A. (2020). Behavioural lateralization in a detour test is not repeatable in fishes. Animal Behaviour, 167, 55–64. https://doi.org/10.1016/j.anbehav.2020.06.025
Staven, F. R., Nordeide, J. T., Imsland, A. K., Andersen, P., Iversen, N. S., & Kristensen, T. (2019). Is habituation measurable in lumpfish Cyclopterus lumpus when used as cleaner fish in atlantic salmon salmo salar aquaculture? Frontiers in Veterinary Science, 6, 1–9. https://doi.org/10.3389/fvets.2019.00227
Versace, E., Morgante, M., Pulina, G., & Vallortigara, G. (2007). Behavioural lateralization in sheep (Ovis aries). Behavioural Brain Research, 184(1), 72–80. https://doi.org/10.1016/j.bbr.2007.06.016