
The biometry of Cardisoma crassum (Brachyura: Gecarcinidae) from two folkloric festivals in Veraguas, Panama
La biometría de Cardisoma crassum (Brachyura: Gecarcinidae) de dos festivales folclóricos en Veraguas, Panamá
Roberto Carlos Lombardo González1,2
https://orcid.org/0000-0002-0279-8621
1Universidad de Panamá. Centro Regional Universitario de Veraguas. Centro de Capacitación, Investigación y Monitoreo de la Biodiversidad (CCIMBIO).
2Sistema Nacional de Investigación de la Secretaría Nacional de Ciencia, Tecnología e Innovación (SNI-SENACYT)
Autor correspondiente: roberto.lombardo@up.ac.pa
Enviado el: 30 de noviembre de 2023 Aceptado el: 2 de enero de 2024
https://doi.org/10.59722/rcvn.v1i2.705
Abstract
This study explores the biometry of Cardisoma crassum individuals presented in two folkloric festivals within the Montijo Gulf in 2022 and 2023. Crabs in the Rio de Jesús (RJ) festival from 2022 exhibited a male biased sex ratio, contrasting Montijo's (MO) even sex ratio in 2023. Carapace widths were similar between festivals (RJ, 65.59 ± 6.56 mm; MO: 64.29 ± 5.92 mm). Sex-based size variations were consistent within years but not between years. Localities showed significant size and sex ratio differences, while transportation distance to the festival venues did not correlate with crab size. Additionally, a comparison between current data and previous studies from 2014–2015, 2021 and 2022, highlighted size differences among sites might not be related to yearly increasing crab demand; instead, site-specific extraction pressure may be an underlying factor. Festivals consumed approximately 198 crab bundles in Rio de Jesús in 2022 (≈ 2,376 individuals) and an estimated 207 bundles in Montijo in 2023 (≈ 2,484 individuals). This study unveils complex dynamics in C. crassum populations at the local level, encompassing sex ratios, size variations, and economic dimensions within festival contexts. Understanding these multifaceted dynamics is crucial for effectively managing these coastal crab populations in the future.
Keywords
Blue crab, cephalothorax, fishery, Montijo, Rio de Jesús.
Resumen
Este estudio analiza la biometría de Cardisoma crassum en dos festivales folclóricos del Golfo de Montijo en 2022 y 2023. En el festival de Río de Jesús (RJ) de 2022, los cangrejos mostraron una marcada tasa de sexos parcializada hacia los machos, a diferencia de Montijo (MO) en 2023, donde la proporción entre sexos fue equilibrada. El ancho del caparazón fue similar (RJ: 65.59 ± 6.56 mm; MO: 64.29 ± 5.92 mm) entre festivales. Las variaciones de talla según el sexo fueron consistentes dentro de cada año, pero no entre años. Se observaron diferencias en talla y proporción de sexos entre localidades, mientras que la distancia de transporte hasta los festivales no estuvo asociada a variación en tallas. La comparación entre datos actuales y 2014-2015, 2021 y 2022, resaltó que diferencias de talla entre sitios podrían no estar relacionadas con un aumento anual en la demanda de cangrejos; en cambio, la presión de extracción específica por sitio podría ser un factor importante. Se consumieron aproximadamente 2,376 cangrejos en Río de Jesús durante 2022 y unos 2,484 en Montijo en 2023. Este estudio revela dinámicas complejas en las poblaciones de C. crassum a nivel local, abordando proporciones de sexos, variaciones en talla y aspectos económicos de festivales. Comprender estas dinámicas será clave para gestionar estas poblaciones en el futuro.
Palabras clave
Cangrejo azul, cefalotórax, pesquería, Montijo, Rio de Jesús.
Introduction
The Blue Crab, Cardisoma crassum, is an integral fishery resource and a prevalent resident of the mangrove ecosystems in the Panamanian Pacific (Lombardo & Rojas, 2022; Vega et al. 2018). Blue crabs are ecosystem engineers (McGaw et al. 2019) that play an essential role in the intricate balance of mangrove ecosystems, through activities such as turning over organic material (Lindquist et al. 2009; Robertson, 1986), and influencing soil chemistry through biological activity (Green, 2004b; Lindquist et al. 2009; Perger et al. 2013; Smith et al. 1991).
This species is captured mainly for subsistence in coastal communities (Hendrickx, 1995) through manual methods and artisanal fishing gear (Vázquez-López & Ramírez-Pérez, 2015; Vega et al. 2018). However, the fishery of C. crassum remains unregulated, leading to uncertainty about its population status. Despite its socioeconomic value for coastal communities (Lombardo & Cedeño, 2023), only three field studies on C. crassum from Panama have been published. Vega et al. (2018) described the artisanal fishery of the blue crab, Lombardo & Rojas (2022) reported the first occurrence of burrow fidelity in C. crassum, while Lombardo & Cedeño (2023) demonstrated size-based segregation; all reports originated from the Montijo Gulf.
Within the gulf, two folkloric festivals occur in October and November: the "Festival del Cangrejo y el Marisco Montijano" in Montijo and the "Festival Mi Ranchito" in Rio de Jesús. These festivals celebrate the blue crab as a cultural and local gastronomic icon, emphasizing its significance as a fishery resource alongside its cultural value (figure 1A‐B). Although these events potentially spike the demand for C. crassum, there exists no formal or continuous record detailing the number of crabs sacrificed, their sizes, or places of origin. The sole published report by Vega et al. (2018) estimates the utilization of approximately 4,800 crabs during 2014 and 2015 in Montijo.
Negative impacts associated with tourism and gastronomic festivals have been documented in other brachyurans and anomurans, like the blue crab Callinectes sapidus (Paolisso, 2007) and the coconut crab Birgus latro (Liu & Lu, 2014), suggesting potential risks for C. crassum. Therefore, understanding the impact of extraction on C. crassum is crucial, as crustacean populations subject to non-industrialized extraction methods can still face overexploitation (João et al. 2022; Lima et al. 2021). Thus, further insight into the potential impact of episodic demand surges for C. crassum is essential for effective future fishery management and conservation (Boenish et al. 2022; Heppell et al. 2005).
Given the typically extended lifespans and delayed reproduction of most gecarcinids (Hartnoll, 1988, 2001; Vogt, 2019), it's plausible that C. crassum is also a slow-growing, long-lived species (Vogt, 2012, 2019) potentially reproducing later in life (Beverton, 1987; Charnov, 1993; Charnov & Berrigan, 1990). This is relevant because intense extraction practices could significantly accelerate population declines in species exhibiting such life history strategies (Beverton, 1987; Heppell et al. 2005; Vogt, 2012). Hence, the potential combination of a slow growth rate and extended lifespan in C. crassum raises concerns about its susceptibility to rapid population declines during yearly demand spikes, such as those occurring during the aforementioned festivals. The current study aims to enhance our understanding of crab biometrics observed during the Montijo and Rio de Jesús festivals. Additionally, it seeks to explore potential trends by comparing records from previous studies in the Montijo Gulf alongside 2022–2023 observations.
Materials and Methods
Study site
According to the “Instituto Nacional de Estadística y Censo” (INEC), in 2023 the total population of Montijo was 6,784 inhabitants, while Rio de Jesús totaled 4,822 residents (INEC, 2023). The Montijo and Rio de Jesús counties extend into the Montijo Gulf, which is also a RAMSAR site of international ecological significance, spanning an area of 808 km2 (RAMSAR, 2023). The area between both counties is irrigated by the San Pablo and San Pedro River watersheds (figure 1C). According to Köppen's classification, the climate within the Montijo Gulf is defined as humid-tropical (Beck et al. 2018). This region is notably characterized by an annual precipitation exceeding 2,500 mm, coupled with temperatures fluctuating between 24.8 and 35.9 °C (Vega et al. 2021).
Sampling protocol
The initial sampling campaign took place in Rio de Jesús in November 2022, while the subsequent sampling occurred in Montijo, October 2023. Unfortunately, the November 2023 edition of the “Mi Ranchito” festival in Rio de Jesús was canceled due to nationwide protests against metallic mining, preventing the collection of data. The sampling process commenced with simple interviews aimed at familiarizing the participants with the study's objectives. These inquiries were directed toward crucial information, such as the estimated number of acquired crabs, their cost, and their origins. In Rio de Jesús, data collection spanned the three-day festival period, from November 18th–20th, 2022. Conversely, in Montijo, data was gathered earlier, from October 20th–22nd, 2023, to circumvent roadblocks and protests against mining, which were anticipated during the actual festival dates a week later.
Biometrics and analysis
Initially, the sex of captive crabs was determined by the shape of the abdomen (Hendrickx, 1995), and the reproductive state of the females (ovigerous or not) was recorded, as well as, the carapace width‐length. A Vernier digital caliper (0.1 mm) was used to measure the width (CW) and length (CL) of the carapace. To avoid resampling, the crabs were temporarily marked with white chalk on the carapace. Chi-square tests were used to compare the proportion of sexes. Variances between groups were tested with Bonnett’s or an F-test, depending on normality (Anderson & Darling, 1952). Differences in biometrics between sexes from the same site were explored using the student’s t-test, while ANOVA was used to test for differences between males and females (four groups) from Rio de Jesús and Montijo. The CW and CL of the crabs were compared between sites, sex, and years. Site comparisons specifically emphasized exploring within-sample variations rather than attempting to draw population-level inferences. This cautious approach considers the uncertainty surrounding whether the samples accurately represent the population's status. The biometry of crabs from sites further away from the festival venues might be less influenced by extraction; thus, regression was applied to assess the relationship between crab size and site of origin distance to the festival venues. To determine such distance, the ruler tool in Google Earth Pro was utilized in its path mode (Google, 2022). This involved tracing the trails, streets and main roads from the crabs' site of origin to their corresponding festival venue, calculating the total kilometers traveled.

Figure 1.
Highlights of folkloric festivals and the study site location. A. Traditional crab bundle showcased at the 'Mi Ranchito' festival in Rio de Jesús, November 2022. B. Folkloric blue crab dance, Rio de Jesús, November 2014. C. Sampling sites at Montijo (17 N 494372.00 m E 883306.00 m N) and Rio de Jesús (17 N 482446.00 m E 882649.00 m N). Scale bar = 20 km (Google, 2022).
To estimate the quantity of crabs arriving to the festivals, as well as to project market prices and profits, informal surveys were conducted among food stand owners and festival organizers. Traditionally, crabs are bundled in groups of 12 (figure 1A). This count served as a reference point to approximate the total number of crabs and calculate profits per festival. This was achieved by multiplying the reported number of bundles and minimum price provided by food stand owners and festival organizers.
Results
Biometry from Rio de Jesús and Montijo festivals
In 2022, a total of 78 C. crassum individuals were measured in Rio de Jesús (49♂, 29♀) with a significantly skewed sex ratio towards males (1.7:1, χ2 = 5.13, P = 0.024). The average CW and standard deviation from Rio de Jesús in 2022 were 65.59 ± 6.56 mm, with a minimum and maximum of 53.13 to 83.42 mm, respectively; the average CL was 54.0 ± 5.48 mm, ranging 44.80–68.93 mm.
In 2023, a total of 116 crabs were measured in Montijo (67♂, 49♀), where no significant difference was found in the sex ratio (1.4:1, χ2 = 2.79, P = 0.095). Subsequently, the average crab CW from Montijo in 2023 was 64.29 ± 5.92 mm (51.58–77.89 mm), while the CL averaged 51.77 ± 6.07 mm (36.64–66.46). In Montijo, specifically from La Caleta, a female carrying eggs measuring 59.32 mm CW was recorded.
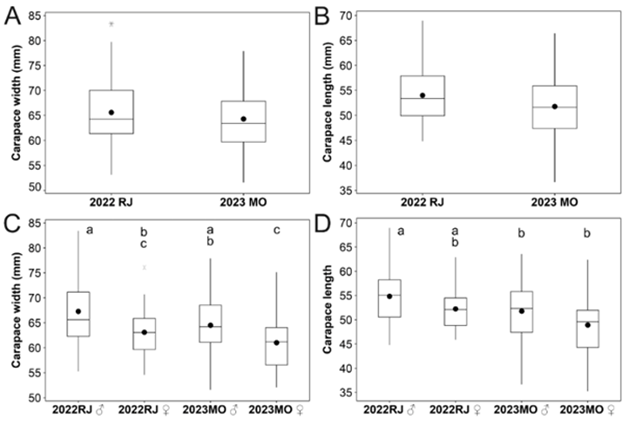
Figure 2.
Cardisoma crassum carapace width and length comparisons between years and the sexes during “Mi Ranchito” (Rio de Jesús = RJ) and "Festival del Cangrejo y el Marisco Montijano" (Montijo = MO) festivals, Veraguas, Panama. Different superscript letters between means show significant differences.
Contrast between festivals
There was no difference in CW between crabs from Rio de Jesús and Montijo festivals (CW, t = 1.44, d.f. = 192, P = 0.151); however, the crabs from Rio de Jesús (2022) were larger in CL (2.23 mm difference) than those from Montijo (2023) (CL, t = 2.61, d.f. = 192, P = 0.010; figure 2A‐B). There was variation between the sexes in CW from Rio de Jesús and Montijo (ANOVA, F(1,3) = 7.54, P < 0.001), where males were larger than the females within years; however, there was no difference between males, nor females across 2022 to 2023 (figure 2C). For CL a different variation pattern emerged (ANOVA, F(1,3) = 7.02, P < 0.001), where Rio de Jesús males from 2022 were larger than Montijo 2023 males and females. Nonetheless, pairwise comparison indicated no difference between the sexes within years from the same site (figure 2D).

Figure 3.
Sites of origin of Cardisoma crassum crabs that arrived at the “Mi Ranchito” (Rio de Jesús, purple) and "Festival del Cangrejo y el Marisco Montijano" (Montijo, yellow) festivals in the Montijo Gulf, Veraguas, Panama. The red square represents the origin (Ponuga) of data from 2021 by (Lombardo & Rojas, 2022). Scale = 10 km (Google, 2022).
Contrasts between sites of origin
The crabs available in the Rio de Jesús festival came from two different sites: Hicaco (46 %) and Las Huacas (54 %). The sample from the Montijo festival contained crabs from three sites: Soná (34 %), La Caleta (40 %) and Puerto Vidal (26 %) (figure 3); see table 1 for a CW‐CL summary.
At the local level the sex ratio was significantly skewed towards male crabs only in the Hicaco site, where the largest male was also captured; the smallest individual, a male, came from Soná. Size differences between the sexes, at the local level, were only detected in crab CW in Las Huacas and Puerto Vidal (table 1).
When male crab CW was compared according to their site of origin, a significant difference was found (ANOVA, F(4,111) = 8.26, P < 0.001); however the pairwise (Tukey) comparisons showed that crabs arriving in Montijo from Soná were smaller, indicating that they were the primary contributor to this observed difference (table 2).
Table 1.
Statistical summary of the carapace width (CW) and length (CL) by site of origin and sexes in Cardisoma crassum from two folkloric festivals in 2022 and 2023 from the Montijo Gulf, Veraguas, Panama. Hicaco (HI), Las Huacas (HU), Soná (SO), La Caleta (CA), Puerto Vidal (VI). SD is the standard
deviation and asterisk represents significance (α = 0.05).
|
UTM coordinates |
Variable |
N |
Mean ± SD |
Min‐Max |
Sex ratio |
Contrast |
|
|
2022 |
17 N 477532.89 m E 844499.88 m N |
CW HI ♂ |
25 |
69.15 ± 8.12 |
55.26 - 83.42 |
2.3:1 χ2 = 5.44 P = 0.020* |
t = 1.83 d.f. = 34 P = 0.076 |
|
CW HI ♀ |
11 |
64.15 ± 5.91 |
54.56 - 76.09 |
||||
|
CL HI ♂ |
25 |
56.63 ± 6.61 |
45.63 - 68.93 |
t = 1.33 d.f. = 34 P = 0.193 |
|||
|
CL HI ♀ |
11 |
53.67 ± 4.89 |
47.46 - 62.91 |
||||
|
17 N 484204.51 m E 871138.31 m N |
CW HU ♂ |
24 |
65.34 ± 4.00 |
59.90 - 74.35 |
1.3:1 χ2 = 0.86 P = 0.355 |
t = 2.19 d.f. = 40 P = 0.034* |
|
|
CW HU ♀ |
18 |
62.46 ± 4.47 |
55.13 - 70.71 |
||||
|
CL HU ♂ |
24 |
52.96 ± 3.98 |
44.80 - 58.25 |
t = 1.30 d.f. = 40 P = 0.201 |
|||
|
CL HU ♀ |
18 |
51.37 ± 3.79 |
45.87 - 58.11 |
||||
|
2023 |
17 N 468271.70 m E 880734.99 m N |
CW SO ♂ |
20 |
60.28 ± 4.63 |
51.58 - 67.85 |
1:1 χ2 = 0.00 P = 1.00 |
t = 1.71 d.f. = 38 P = 0.095 |
|
CW SO ♀ |
20 |
57.74 ± 4.76 |
52.07 - 68.03 |
||||
|
CL SO ♂ |
20 |
48.86 ± 5.64 |
41.65 - 64.44 |
t = 1.32 d.f. = 38 P = 0.193 |
|||
|
CL SO ♀ |
20 |
46.40 ± 6.11 |
35.28 - 59.28 |
||||
|
17 N 490841.05 m E 875650.74 m N |
CW CA ♂ |
28 |
66.42 ± 5.19 |
56.84 - 75.41 |
1.6:1 χ2 = 2.17 P = 0.140 |
t = 1.18 d.f. = 44 P = 0.246 |
|
|
CW CA ♀ |
18 |
64.59 ± 5.07 |
59.26 - 75.15 |
||||
|
CL CA ♂ |
28 |
53.11 ± 5.90 |
36.64 - 61.40 |
t = 1.27 d.f. = 44 P = 0.212 |
|||
|
CL CA ♀ |
18 |
50.90 ± 5.60 |
41.93 - 62.40 |
||||
|
17 P 433749.28 m E 886554.80 m N |
CW VI ♂ |
19 |
69.02 ± 5.64 |
58.30 - 77.89 |
1.7:1 χ2 = 2.13 P = 0.144 |
t = 2.03 d.f. = 28 P = 0.052* |
|
|
CW VI ♀ |
11 |
64.78 ± 5.26 |
54.97 - 75.72 |
||||
|
CL VI ♂ |
19 |
55.37 ± 4.74 |
43.20 - 63.56 |
t = 0.57 d.f. = 28 P = 0.573 |
|||
|
CL VI ♀ |
11 |
54.19 ± 6.62 |
42.51 - 66.46 |
Male CW from Hicaco, La Huaca, La Caleta and Puerto Vidal were not significantly different; the same pattern was detected in the females (ANOVA, F(4,73) = 6.14, P < 0.001). Male CL showed significant variation among sites of origin (ANOVA, F(4,111) = 6.21, P < 0.001); however, such variation was only significant in two pairwise comparisons against Soná: Hicaco and Puerto Vidal. Both male and female CL from Soná were smaller than the rest of the sites. For female CL (ANOVA, F(4,73) = 5.14, P = 0.001) only three pairwise comparisons resulted significant against Soná: Hicaco, Las Huacas and Puerto Vidal (table 2).
Crabs from Puerto Vidal were transported an estimated 124.7 Km to Montijo, while crabs from Hicaco were shipped an estimate of 88.6 Km to Rio de Jesús; however no significant relationship was found between distance and average crab size (r2 = 0.242, F(1,4) = 0.96, P = 0.400).
Table 2.
Analysis of variance summary of carapace width (CW) between sites of origin for Cardisoma crassum in two folkloric festivals including 2021 data (Lombardo & Rojas, 2022) from Ponuga. Hicaco (HI), Las Huacas (HU), Soná (SO), La Caleta (CA), Puerto Vidal (VI), Ponuga (PA). SD is the standard deviation and asterisk represents significance.
|
Year |
CW |
N |
Mean ± SD |
Contrast |
Difference |
Tukey (α = 0.05) |
|
2021 |
PA |
164 |
62.69 ± 6.07 |
HI - PA |
4.93 |
t = 4.56, P < 0.001* |
|
HU - PA |
1.17 |
t = 1.15, P = 0.859 |
||||
|
SO - PA |
-3.58 |
t = -3.42, P = 0.008* |
||||
|
CA - PA |
3.02 |
t = 3.08, P = 0.025* |
||||
|
VI - PA |
4.78 |
t = 4.10, P = 0.001* |
||||
|
2022 |
HI |
36 |
67.62 ± 7.79 |
HU - HI |
-3.76 |
t = -2.82, P = 0.054* |
|
SO - HI |
-8.52 |
t = -6.27, P < 0.001* |
||||
|
CA - HI |
-1.91 |
t = -1.46, P = 0.687 |
||||
|
VI - HI |
-0.15 |
t = -0.10, P = 1.000 |
||||
|
HU |
42 |
63.86 ± 4.71 |
SO - HU |
-4.75 |
t = -3.64, P = 0.004* |
|
|
CA - HU |
1.85 |
t = 1.48, P = 0.680 |
||||
|
VI - HU |
3.61 |
t = 2.57, P = 0.104 |
||||
|
2023 |
SO |
39 |
59.10 ± 4.83 |
CA - SO |
6.6 |
t = 5.16, P < 0.001* |
|
VI - SO |
8.37 |
t = 5.86, P < 0.001* |
||||
|
CA |
46 |
65.71 ± 5.17 |
VI - CA |
1.76 |
t = 1.28, P = 0.797 |
|
|
VI |
30 |
67.47 ± 5.80 |
In either festival, around 18–20 food stands served the public, and according to the owners, each stand procures approximately 10 crab bundles, while the organizing committee also purchases ≈ 30 bundles; each containing 12 crabs. During 2022, in Rio de Jesús, an estimate 198 crab bundles were acquired for the festival, totaling ≈ 2,376 individuals. Depending on the size of crabs in the bundle, the local prize varied widely from 8,10,14 up to 20 USD; thus, a precise estimate of profit was not possible. However, if a bundle cost ≈ 8 USD (minimum), then extractors/resellers were estimated to profit at least 1,584 USD. In 2023, it is estimated 207 crab bundles were consumed in the Montijo festival, totaling ≈ 2,484 individuals; thus, by a similar price metric, extractors and/or resellers made at least 1,656 USD in profit.
Discussion
The earliest published record (Vega et al. 2018) of crab statistics from the Montijo Gulf festivals dates back to 2014–2015. Inconsistency of records resulted in an unfortunate eight-year gap (2015–2023) in data on the number and biometry of crabs captured for the festivals, and in general within the gulf. According to festival organizers, from 2007 to 2013, the number of crabs used each year in Montijo varied at around 2,400. In 2014, organizers reported to Vega et al. (2018) the acquisition of approximately 3,000 crabs, while in 2015, organizers bought roughly 1,800 individuals. Comparatively, total crab estimates in the festival editions hint at a steady demand with mild variability throughout the years. Considering that the Rio de Jesús (≈ 2,376 ind. In 2022) and Montijo (≈ 2,484 ind. In 2023) festivals occur in a year, the populations of C. crassum in the gulf might face an approximate annual loss of 4,860 crabs due to fishing mortality related to the festivals. At present, the correlation between the annual demand for crab and its impact on the population of C. crassum remains uncertain. However, it's conceivable that the extraction process could affect this species, leading to potential consequences such as diminished size structure, altered sex ratios, and reduced fecundity (Beverton, 1987; Pauly & Froese, 2014; Sato & Goshima, 2006). Similar effects resulting from extraction have been reported in other land crab species, such as Birgus latro (Sato & Yoseda, 2010) and Ucides cordatus (Diele et al., 2005), suggesting a possibility of similar repercussions for C. crassum.
If the effect of festival demand is substantial, then the size of crabs is expected to decrease as extraction is biased towards larger sizes; usually males (Govender et al. 2008; Hartnoll et al. 2006).
Hypothetically, impacted populations would suffer size structure shifts, where homogeneity in size between the sexes is expected. In the Montijo Gulf, typical carapace width difference between the sexes has been observed at the regional level, where males were larger than females (Lombardo & Cedeño, 2023; Lombardo & Rojas, 2022; Vega et al. 2018). Interestingly, such shift towards homogeneity in size structure was not observed in the present study; there was variation in carapace width between the sexes during 2022 in Rio de Jesús and 2023 in Montijo, with males being consistently larger than females. Thus, large size extraction effects are still below critical significance.
Furthermore, there were no discernible differences among males or females across 2022 and 2023, and in general, no significant reduction in carapace width was noted between the crabs from the festivals in 2022 and 2023. These results are in contrast to the expected effect of a hypothetical high crab demand; however, the results must be interpreted with caution, as it is difficult to rule out demand from festivals, mainly because the samples might not be representative of each site considered. However, when compiling the data between years, a decrease in crab size (2021-2023) was not detected. Larger average carapace width from 2014-2015 (Vega et al. 2018) and 2021 (Lombardo & Rojas, 2022) might be expected as well, but they do not square evenly with large crabs from 2022-2023. In this case, the crabs from 2021 (Ponuga) were significantly smaller than those from 2022-2023 (see Table 2). This local discrepancy through time implies the size structure of C. crassum might not be influenced by an increasing year-to-year extraction pressure. Alternatively, a future direction for research into this topic would assess the concentrated, site-specific extraction impact, which could have a significant effect in shaping the size structure of C. crassum, as shown in the carapace width comparisons by site. Determining correlates of accessibility, habitat degradation, land use and C. crassum size structure might yield interesting results in the future.
Crabs in the Rio de Jesús festival (2022) exhibited a male skewed sex ratio, while the sex ratio in Montijo (2023) was even. The sex ratio of C. crassum within the Montijo Gulf has been reported to be male biased in 2014 and 2015 (Vega et al., 2018), even in 2021 (Lombardo & Rojas, 2022), and male biased in 2022 (Lombardo& Cedeño, 2023). Normally, the sex ratio in brachyuran species tends to skew towards males (Wenner, 1972). Variation in size and sex ratios may be explained by extraction effects, unequal growth due to divergent reproductive strategies between the sexes, variable maximum sizes, and lifespan (Diele et al. 2005; Ewers-Saucedo, 2019; Wenner, 1972). Female skewed sex ratios are often interpreted as another sign of extraction impact or habitat degradation (Pardo et al. 2017; Quiñones-Llópiz et al. 2021). In the region, sex ratios seem to vary between sites, but also seem to be independent of the year, suggesting possible differences in the sampling effort or extraction activity associated to particular sites; nonetheless, it is worth noting that to our knowledge, no study in Panama has reported a female biased sex ratio for C. crassum. This suggests, perhaps at the local level, populations might retain functional recovery capacity (Ewers-Saucedo, 2019; Wenner, 1972).
Among the crabs already present at the festival venues, we observed a gravid female, while the rest fell within the carapace width range (47.11–70.90 mm) documented for egg-carrying female crabs in Vega et al. (2018), Lombardo & Rojas (2022), and Lombardo & Cedeño (2023). A similar range was reported in Mexico (Vargas-Téllez & Vázquez-López, 2016) for ovigerous C. crassum females, between 46.40 and 71.48 mm, with an average of 59.7±6.2 mm. These ranges for maturity are relevant because in C. crassum the carapace width is correlated with fecundity (Vargas-Téllez & Vázquez-López, 2016). Although more work remains to be done to confirm the age and size at first maturity, the maximum and average sizes in the present study suggest that a proportion of the females arriving at the festivals might be early breeding females. This has important implications concerning crab demand, in which fishing mortality has been shown to reduce individual size and consequently female reproductive potential (Hines et al., 2003; Hjelset, 2014; Jivoff, 2003).
It is unknown how long does it take for C. crassum to reach the minimum size for first maturity (46.40–47.11 mm? CW). Nevertheless, considering the life history of gecarcinids, high extraction pressure at the local level could have severe impacts as this group of land crabs is slow growing and long lived, reproducing later in life. For example, Gecarcinus lateralis and Gecarcinus quadratus, have expected lifespans of 10 years; Cardisoma armatum, is reported to live for 12 years (Rademacher & Mengedoht, 2011); Gecarcinus ruricola, has a lifetime of 15 years (Hartnoll et al. 2006); Cardisoma guanhumi (Wolcott, 1988) and Gecarcoidea natalis, (Green, 2004a; Linton & Greenaway, 1997; Wolcott, 1988) have estimated lifespans of 20 years (Vogt, 2012, 2019). This suggests there might be a risk for the recovery potential of overfished C. crassum stocks if extraction remains unregulated (Ewers-Saucedo, 2019; Hines et al. 2003; Jivoff, 2003; Sato & Goshima, 2006).
Conclusions
· Cardisoma crassum holds significant importance as a fishery resource for coastal communities as demonstrated by the generation of profits for extractors and resellers throughout the years. It is also a celebrated figure in folkloric festivals highlighting its cultural value.
· Differences in crab size might be better explained by extraction effort associated with particular sites, not yearly increases in crab demand.
· No significant reduction in crab size was detected between festival years, contrary to the expected effects of high crab demand.
· Despite variations in sex ratios across different sites in the Montijo Gulf, the consistency in the absence of a female-biased ratio throughout multiple years suggests a level of resilience within the local populations.
· The observed crab carapace widths at festivals indicate potential prior maturity for all females, with indications of early breeding females. This raises concerns about fishing mortality affecting both individual size and future female reproductive potential.
Acknowledgments
We are grateful to Luz Maria Batista for her cooperation in meeting the food stand owners as well as Enrique Garrido, Nelly Reyes and José Batista for access to their crabs in Montijo. We also thank the owners of the restaurant El Gabriel in Rio de Jesús for access to their crabs.
References
Anderson, T. W., & Darling, D. A. (1952). Asymptotic theory of certain “goodness of fit” criteria based on stochastic processes. Annals of Mathematical Statistics, 23(2), 193–212. https://doi.org/10.1214/AOMS/1177729437
Beck, H. E., Zimmermann, N. E., McVicar, T. R., Vergopolan, N., Berg, A., & Wood, E. F. (2018). Present and future köppen-geiger climate classification maps at 1-km resolution. Scientific Data, 5. https://doi.org/10.1038/sdata.2018.214
Beverton, R. J. H. (1987). Longevity in fish: Some ecological and evolutionary considerations. In A. D. Woodhead & K. H. Thompson (Eds.), Evolution of Longevity in Animals. Springer. https://doi.org/10.1007/978-1-4613-1939-9_12
Boenish, R., Kritzer, J. P., Kleisner, K., Steneck, R. S., Werner, K. M., Zhu, W., Schram, F., Rader, D., Cheung, W., Ingles, J., Tian, Y., & Mimikakis, J. (2022). The global rise of crustacean fisheries. Frontiers in Ecology and the Environment, 20(2), 102–110. https://doi.org/10.1002/FEE.2431
Charnov, E. L. (1993). Life history invariants: Some explorations of symmetry in evolutionary ecology (1st ed.). Oxford University Press.
Charnov, E. L., & Berrigan, D. (1990). Dimensionless numbers and life history evolution: Age of maturity versus the adult lifespan. Evolutionary Ecology, 4(3), 273–275. https://doi.org/10.1007/BF02214335/METRICS
Diele, K., Koch, V., & Saint-Paul, U. (2005). Population structure, catch composition and CPUE of the artisanally harvested mangrove crab Ucides cordatus (Ocypodidae) in the Caeté estuary, North Brazil: Indications for overfishing? Aquatic Living Resources, 18(2), 169–178.
Ewers-Saucedo, C. (2019). Evaluating reasons for biased sex ratios in Crustacea. Invertebrate Reproduction & Development, 63(3), 222–230. https://doi.org/10.1080/07924259.2019.1588792
Google. (2022). Google Earth Pro (7.3.6.9345 (64-bit)). Google.
Govender, Y., Sabat, A. M., & Cuevas, E. (2008). Effects of land-use/land-cover changes on land crab, Cardisoma guanhumi, abundance in Puerto Rico. Journal of Tropical Ecology, 24(4), 417–423. https://doi.org/10.1017/S0266467408005130
Green, P. T. (2004a). Burrow dynamics of the red land crab Gecarcoidea natalis (Brachyura, Gecarcinidae) in rain forest on Christmas Island (Indian Ocean). Journal of Crustacean Biology, 24(2), 340–349.
Green, P. T. (2004b). Field observations of moulting and moult increment in the Red Land Crab, Gecarcoidea natalis (Brachyura, Gecarcinidae), on Christmas Island (Indian Ocean). Crustaceana, 77(1), 125–128. https://doi.org/10.1163/156854004323037937
Hartnoll, R. G. (1988). Growth and Molting. In W. Burggren & B. McMahon (Eds.), Biology of the Land Crabs (pp. 186–210). Cambridge University Press. https://doi.org/10.1017/CBO9780511753428.007
Hartnoll, R. G. (2001). Growth in Crustacea - Twenty years on. Hydrobiologia, 449(1), 111–122. https://doi.org/10.1023/A:1017597104367/METRICS
Hartnoll, R. G., Baine, M. S. P., Grandas, Y., James, J., & Atkin, H. (2006). Population Biology of the Black Land Crab, Gecarcinus Ruricola, in the San Andres Archipelago, Western Caribbean. Journal of Crustacean Biology, 26(3), 316–325. https://doi.org/10.1651/C-2640.1
Hendrickx, M. E. (1995). Cangrejos. In Guía FAO para la identificación de especies para los fines de la pesca: Pacifico Centro-Oriental. (1st ed., Vol. 1, pp. 581–636). FAO. 646.
Heppell, S. S., Heppell, S. A., Read, A. J., & Crowder, L. B. (2005). Effects of fishing on long-lived marine organisms. In E. A. Norse & L. B. Crowder (Eds.), Marine conservation biology: The science of maintaining the sea’s biodiversity (pp. 211–231). Island Press.
Hines, A. H., Jivoff, P. R., Bushmann, P. J., Van Montfrans, J., Reed, S. A., Wolcott, D. L., & Wolcott, T. G. (2003). Evidence for sperm limitation in the blue crab, Callinectes sapidus. Bulletin of Marine Science, 72(2), 287–310.
Hjelset, A. M. (2014). Fishery-induced changes in Norwegian red king crab (Paralithodes camtschaticus) reproductive potential. ICES Journal of Marine Science, 71(2), 365–373. https://doi.org/10.1093/ICESJMS/FST126
INEC. (2023, October 9). Contraloría General de la República de Panamá - Instituto Nacional de Estadística y Censo. Resultados Finales Básicos XII Censo Nacional de Población y VIII de Vivienda 2023. https://www.inec.gob.pa/publicaciones/Default2.aspID_CATEGORIA=19&ID_SUBCATEGORIA=71
Jivoff, P. (2003). A review of male mating success in the blue crab, Callinectes sapidus, in reference to the potential for fisheries-induced sperm limitation. Bulletin of Marine Science, 72(2), 273–286.João, M. C. A., Duarte, R. C., da Silva, L. S. B., Freire, A. S., & Pinheiro, M. A. A. (2022). Sexual maturity of an endemic insular land crab: Priority information toward the conservation of, Johngarthia lagostoma. The Biological Bulletin, 243(1), 14–27. https://doi.org/10.1086/720581
Lima, M. C., Pereira, C. A. M., Araújo, M. S. L. C., Rodrigues, G. G., & Nicacio, G. (2021). Seasonal variation in biometric parameters in a population of the endangered blue land crab (Cardisoma guanhumi): Indicators for assessment and management. Regional Studies in Marine Science, 45, 101804. https://doi.org/10.1016/J.RSMA.2021.101804
Lindquist, E. S., Krauss, K. W., Green, P. T., O’Dowd, D. J., Sherman, P. M., & Smith, T. J. (2009). Land crabs as key drivers in tropical coastal forest recruitment. Biological Reviews, 84(2), 203–223.
Linton, S. M., & Greenaway, P. (1997). Urate deposits in the gecarcinid land crab Gecarcoidea natalis are synthesised de novo from Excess dietary nitrogen. Journal of Experimental Biology, 200(17), 2347–2354. https://doi.org/10.1242/JEB.200.17.2347
Liu, T. M., & Lu, D. J. (2014). The cultural and ecological impacts of aboriginal tourism: A case study on Taiwan’s Tao tribe. SpringerPlus, 3(1), 1–9. https://doi.org/10.1186/2193-1801-3-347/METRICS
Lombardo, R. C., & Cedeño, L. M. (2023). Segregation by size in the mouthless crab Cardisoma crassum (Brachyura: Gecarcinidae) from Ponuga, Veraguas, Panama. Revista Ciencias Marinas y Costeras, 15(1), 27–44. https://doi.org/10.15359/revmar.15-1.2
Lombardo, R. C., & Rojas, M. (2022). Burrow fidelity in the blue crab, Cardisoma crassum Smith, 1870 (Brachyura: Gecarcinidae) from the Ponuga River, Veraguas, Panama. Nauplius, 30, e2022033. https://doi.org/10.1590/2358-2936e2022033
McGaw, I. J., Van Leeuwen, T. E., Trehern, R. H., & Bates, A. E. (2019). Changes in precipitation may alter food preference in an ecosystem engineer, the black land crab, Gecarcinus ruricola. PeerJ, 2019(5), 1–28. https://doi.org/10.7717/peerj.6818
Paolisso, M. (2007). Taste the traditions: Crabs, crab cakes, and the Chesapeake Bay blue crab fishery. American Anthropologist, 109(4), 654–665. https://doi.org/10.1525/AA.2007.109.4.654
Pardo, L. M., Riveros, M. P., Fuentes, J. P., Pinochet, R., Cárdenas, C., & Sainte-Marie, B. (2017). High fishing intensity reduces females’ sperm reserve and brood fecundity in a eubrachyuran crab subject to sex- and size-biased harvest. ICES Journal of Marine Science, 74(9), 2459–2469. https://doi.org/10.1093/ICESJMS/FSX077
Pauly, D., & Froese, R. (2014). Fisheries Management. In Encyclopedia of Life Sciences. John Wiley & Sons, Ltd. https://doi.org/10.1002/9780470015902.A0003252.PUB3
Perger, R., Cortés, J., & Pacheco, C. (2013). Closing a distributional gap of over 3000 km and encountering an invisible barrier: New presence/absence data for Johngarthia planata Stimpson, 1860 (Decapoda,
Brachyura, Gecarcinidae) for Central America and biogeographic notes on East Pacific Gecarcinidae. Crustaceana, 86(3), 268–277. https://doi.org/10.1163/15685403-00003172
Quiñones-Llópiz, J.-D., Rodríguez-Fourquet, C., Luppi, T., Farias, N.-E., Quiñones-Llópiz, J.-D., Rodríguez-Fourquet, C., Luppi, T., & Farias, N.-E. (2021). Size distribution and sex ratio between populations of the artisanal harvested land crab Cardisoma guanhumi (Decapoda: Gecarcinidae), with the estimation of relative growth and size at sexual maturity in Puerto Rico. Revista de Biología Tropical, 69(3), 989–1003. https://doi.org/10.15517/RBT.V69I3.45570
Rademacher, M., & Mengedoht, O. (2011). Krabben-Fibel (U. Wesollek-Rottmann, Ed.; 1st ed., Vol. 1). Dähne Verlag.
RAMSAR. (2023). The List of Wetlands of International Importance. Ramsar List. https://www.ramsar.org/document/list-wetlands-international-importance-ramsar-list
Robertson, A. I. (1986). Leaf-burying crabs: Their influence on energy flow and export from mixed mangrove forests (Rhizophora spp.) in northeastern Australia. Journal of Experimental Marine Biology and Ecology, 102(2–3), 237–248. https://doi.org/10.1016/0022-0981(86)90179-6
Sato, T., & Goshima, S. (2006). Impacts of male-only fishing and sperm limitation in manipulated populations of an unfished crab, Hapalogaster dentata. Marine Ecology Progress Series, 313, 193–204. https://doi.org/10.3354/MEPS313193
Sato, T., & Yoseda, K. (2010). Influence of size- and sex-biased harvesting on reproduction of the coconut crab Birgus latro. Marine Ecology Progress Series, 402, 171–178. https://doi.org/10.3354/MEPS08369
Smith, T. J., Boto, K. G., Frusher, S. D., & Giddins, R. L. (1991). Keystone species and mangrove forest dynamics: the influence of burrowing by crabs on soil nutrient status and forest productivity. Estuarine, Coastal and Shelf Science, 33(5), 419–432. https://doi.org/10.1016/0272-7714(91)90081-L
Vargas-Téllez, I., & Vázquez-López, H. (2016). Fecundity of Cardisoma crassum Smith, 1870 in an estuary of the Mexican Pacific. Biología, Ciencia y Tecnología, 9(35), 632–641.
Vázquez-López, H., & Ramírez-Pérez, T. (2015). Aspects of growth in the terrestrial crab Cardisoma crassum Smith, 1870 (Crustacea: Brachyura: Gecarcinidae) from El Salado estuary, Puerto Vallarta, Jalisco, México. Mitteilungen Klosterneuburg, 65(2), 82–99.
Vega, A. J., Mena, C., & Robles, Y. (2018). Pesca artesanal Cardisoma crassum (Cangrejo) y Ucides occidentalis (Mangote) en el sector nororiental del Golfo de Montijo, Pacifico de Panamá. 20, 5–22.
Vega, A. J., Robles, Y., Alvarado, O., & Cedeño, C. (2021). Estructura de tallas, distribución y abundancia de Anadara tuberculosa (Bivalvia: Arcidae) en dos sistemas de manglar del Pacífico de Panamá. Revista de Biología Tropical, 69(2), 422–433. https://doi.org/10.15517/RBT.V69I2.43934
Vogt, G. (2012). Ageing and longevity in the Decapoda (Crustacea): A review. Zoologischer Anzeiger - A Journal of Comparative Zoology, 251(1), 1–25. https://doi.org/10.1016/J.JCZ.2011.05.003
Vogt, G. (2019). A compilation of longevity data in decapod crustaceans. Nauplius, 27(e2019011). https://doi.org/10.1590/2358-2936e2019011
Wenner, A. M. (1972). Sex ratio as a function of size in marine Crustacea. The American Naturalist, 106(949), 321–350.
Wolcott, T. G. (1988). Ecology. In W. Burggren & B. McMahon (Eds.), Biology of the Land Crabs (pp. 55–95). Cambridge University Press. https://doi.org/doi:10.1017/CBO9780511753428.004